En los procesos catabólicos las moléculas orgánicas se van degradando, paso a paso, hasta formar otras moléculas más simples y, finalmente, sustancias inorgánicas. Globalmente son procesos de oxidación en los que las moléculas orgánicas van perdiendo electrones que, tras pasar por una cadena transportadora, son captados por una molécula aceptora de electrones final. En esos procesos la energía liberada permite la formación de moléculas de ATP.
Según quien sea el aceptor final de electrones se pueden diferenciar dos modalidades:
§ Fermentación. El aceptor final de electrones es un compuesto orgánico, por lo que se trata de una oxidación incompleta y un proceso anaerobio.
§ Respiración celular. El aceptor final de electrones es una sustancia inorgánica.
Si es el O2, se trata de una respiración aerobia que realizan la mayoría de los organismos; si es otro compuesto inorgánico (NO3-, SO42-, CO2), se trata de una respiración anaerobia, exclusiva de ciertos microorganismos.
Los procesos catabólicos se pueden iniciar a partir de varios sustratos orgánicos, especialmente glúcidos, lípidos y proteínas. En cualquier caso, al final, las diferentes rutas metabólicas de la respiración celular confluyen en el ciclo de Krebs, y los electrones liberados en el proceso pasan a la cadena respiratoria para formar ATP por fosforilación oxidativa. Una variante a este esquema general es el desvío o la ruta alternativa de las fermentaciones.
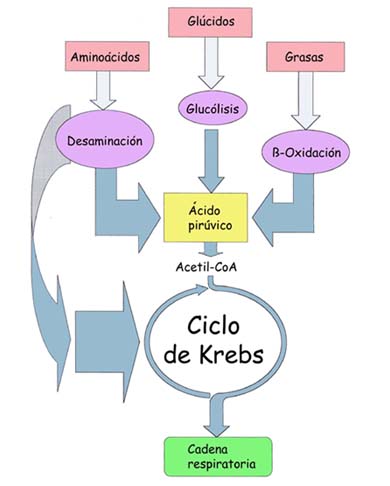
RESPIRACIÓN CELULAR
El proceso por el cual las células degradan las moléculas de alimento para obtener energía recibe el nombre de respiración celular.
La respiración celular es una reacción exergónica, donde parte de la energía contenida en las moléculas de alimento es utilizada por la célula para sintetizar ATP. Decimos parte de la energía porque no toda es utilizada, sino que una parte se pierde.
Aproximadamente el 40% de la energía libre emitida por la oxidación de la glucosa se conserva en forma de ATP. Cerca del 75% de la energía de la nafta se pierde como calor de un auto; solo el 25% se convierte en formas útiles de energía. La célula es mucho más eficiente.
La respiración celular es una combustión biológica y puede compararse con la combustión de carbón, bencina, leña. En ambos casos moléculas ricas en energía son degradadas a moléculas más sencillas con la consiguiente liberación de energía.
Tanto la respiración como la combustión son reacciones exergónicas.
Sin embargo existen importantes diferencias entre ambos procesos. En primer lugar la combustión es un fenómeno incontrolado en el que todos los enlaces químicos se rompen al mismo tiempo y liberan la energía en forma súbita; por el contrarío la respiración es la degradación del alimento con la liberación paulatina de energía. Este control está ejercido por enzimas específicas.
En segundo lugar la combustión produce calor y algo de luz. Este proceso transforma energía química en calórica y luminosa. En cambio la energía liberada durante la respiración es utilizada fundamentalmente para la formación de nuevos enlaces químicos (ATP).
La respiración celular puede ser considerada como una serie de reacciones de óxido-reducción en las cuales las moléculas combustibles son paulatinamente oxidadas y degradadas liberando energía. Los protones perdidos por el alimento son captados por coenzímas.
La respiración ocurre en distintas estructuras celulares. La primera de ellas es la glucólisis que ocurre en el citoplasma. La segunda etapa dependerá de la presencia o ausencia de O2 en el medio, determinando en el primer caso la respiración aeróbica (ocurre en las mitocondrias), y en el segundo caso la respiración anaeróbica o fermentación (ocurre en el citoplasma).

GLUCÓLISIS
La glucólisis, tiene lugar en una serie de nueve reacciones, cada una catalizada por una enzima específica, hasta formar dos moléculas de ácido pirúvico, con la producción concomitante de ATP. La ganancia neta es de dos moléculas de ATP, y dos de NADH por cada molécula de glucosa.
Las reacciones de la glucólisis se realizan en el citoplasma, como ya adelantáramos y pueden darse en condiciones anaerobias; es decir en ausencia de oxígeno.
Los primeros cuatro pasos de la glucólisis sirven para fosforilar (incorporar fosfatos) a la glucosa y convertirla en dos moléculas del compuesto de 3 carbonos gliceraldehído fosfato (PGAL). En estas reacciones se invierten dos moléculas de ATP a fin de activar la molécula de glucosa y prepararla para su ruptura.
Paso 1
La serie de reacciones glucolíticas se inicia con la activación de la glucosa
Glucosa + ATP
La reacción del ATP con la glucosa para producir glucosa 6-fosfatoy ADP es exergónica. Parte de la energía liberada se conserva en el enlace que une al fosfato con la molécula de glucosa que entonces se energiza.
Paso 2
La glucosa 6-fosfato sufre una reacción de reordenamiento catalizada por una isomerasa, con lo que se forma fructosa 6-fosfato.
Paso 3
La fructosa 6-fosfato acepta un segundo fosfato del ATP, con lo que se genera fructosa 1,6-difosfato; es decir fructosa con fosfatos en las posicio-nes 1 y 6.
La enzima que regula esta reacción es la fosfofructocinasa.
Nótese que hasta ahora se han invertido dos moléculas de ATP y no se ha recuperado energía.
La fosfofructocinasa es una enzima alostérica, el ATP es un efector alostérico que la inhibe. La interacción alostérica entre ellos es el principal mecanismo regulador de la glucólisis. Si existe ATP en cantidades suficientes para otros fines de la célula, el ATP inhibe la actividad de la enzima y así cesa la producción de ATP y se conserva glucosa. Al agotar la célula la provisión de ATP, la enzima se desinhibe y se reanuda la degradación de la glucosa. Este es uno de los puntos principales del control de la producción de ATP.
Paso 4
La fructosa 1,6 -difosfato se divide luego en dos azúcares de 3 carbonos, gliceraldehído 3-fosfato y dihidroxiacetona fosfato. La dihidroxiacetona fosfato es convertida enzimáticamente (isomerasa) en gliceraldehído fósfato. Todos los pasos siguientes deben contarse dos veces para tener en cuenta el destino de una molécula de glucosa.
Debemos recordar que hasta el momento no se ha obtenido ninguna energía biológicamente útil. En reacciones subsecuentes, la célula recupera parte de la energía contenida en el PGAL.
Paso 5
Las moléculas de PGAL se oxidan es decir, se eliminan átomos de hidrógeno con sus electrones, y el NAD+ se reduce a NADH. Esta es la primera reacción de la cual la célula cosecha energía. El producto de esta reacción es el fosfoglicerato. Este compuesto reacciona con un fosfato inorgánico (Pi) para formar 1,3 difosfoglicerato. El grupo fosfato recién incorporado se encuentra unido por medio de un enlace de alta energía.
Paso 6
El fosfato rico en energía reacciona con el ADP para formar ATP. (en total dos moléculas de ATP por molécula de glucosa). Esa transferencia de energía desde un compuesto con un fosfato, de alta energía se conoce como fosforfiación.
Paso 7
El grupo fosfato remanente se transfiere enzimáticamente de la posición 3 a la posición 2 (ácido 2-fosfoglicérico).
Paso 8
En este paso se elimina una molécula de agua del compuesto 3 carbono. Este reordenamiento interno de la molécula concentra energía en la vecindad del grupo fosfato. El producto es el ácido fosfoenolpirúvico (PEP).
Paso 9
El ácido fosfoenolpirúvico tiene la capacidad de transferir su grupo fosfato a una molécula de ADP para formar ATP y ácido pirúvico. (dos moléculas de ATP y ácido pirúvico por cada molécula de glucosa).

LA FERMENTACIÓN.
Además de la respiración otra forma anaeróbica de degradar la glucosa y de producir energía es la fermentación. La fermentación es la degradación de glucosa y liberación de energía utilizando sustancias orgánicas, aceptadores finales de electrones. La fermentación es una “medida de emergencia” para producir oxígeno cuando ésta escasea. La primera parte de la fermentación es la glucólisis. Las células de levadura llevan a cabo la fermentación que produce alcohol etílico y CO2 la cual se conoce como fermentación alcohólica
La fermentación es un proceso catabólico de oxidación incompleta, totalmente anaeróbico, siendo el producto final un compuesto orgánico. Estos productos finales son los que caracterizan los diversos tipos de fermentaciones.
Fue descubierta por Louis Pasteur, que la describió como la vie sans l´air (la vida sin el aire). La fermentación típica es llevada a cabo por las levaduras. También algunos metazoos y protistas son capaces de realizarla.
El proceso de fermentación es anaeróbico ya que se produce en ausencia de oxígeno; ello significa que el aceptor final de los electrones del NADH producido en la glucólisis no es el oxígeno, sino un compuesto orgánico que se reducirá para poder reoxidar el NADH a NAD+. El compuesto orgánico que se reduce (acetaldehído, piruvato, ...) es un derivado del sustrato que se ha oxidado anteriormente.
En los seres vivos, la fermentación es un proceso anaeróbico y en él no interviene la mitocondria ni la cadena respiratoria. Son propias de los microorganismos, como algunas bacterias y levaduras. También se produce la fermentación en la mayoría de las células de los animales (incluido el hombre), excepto en las neuronas que mueren rápidamente si no pueden realizar la respiración celular; algunas células, como los eritrocitos, carecen de mitocondrias y se ven obligadas a fermentar; el tejido muscular de los animales realiza la fermentación láctica cuando el aporte de oxígeno a las células musculares no es suficiente para el metabolismo aerobio y la contracción muscular.
Desde el punto de vista energético, las fermentaciones son muy poco rentables si se comparan con la respiración aerobia, ya que a partir de una molécula de glucosa sólo se obtienen 2 moléculas de ATP, mientras que en la respiración se producen 36. Esto se debe a la oxidación del NADH, que en lugar de penetrar en la cadena respiratoria, cede sus electrones a compuestos orgánicos con poco poder oxidante.
En la industria la fermentación puede ser oxidativa, es decir, en presencia de oxígeno, pero es una oxidación aeróbica incompleta, como la producción de ácido acético a partir de etanol.
Las fermentaciones pueden ser: naturales, cuando las condiciones ambientales permiten la interacción de los microorganismos y los sustratos orgánicos susceptibles; o artificiales, cuando el hombre propicia condiciones y el contacto referido

El ciclo de Krebs
Reacción 1: Citrato sintasa (De oxalacetato a citrato)
Reacción 2: Aconitasa (De citrato a isocitrato)
Reacción 3: Isocitrato deshidrogenasa (De isocitrato a oxoglutarato)
Reacción 4: α-cetoglutarato deshidrogenasa (De oxoglutarato a Succinil-CoA)
Reacción 5: Succinil-CoA sintetasa (De Succinil-CoA a succinato)
Reacción 6: Succinato deshidrogenasa (De succinato a fumarato)
Reacción 7: Fumarasa (De fumarato a L-malato)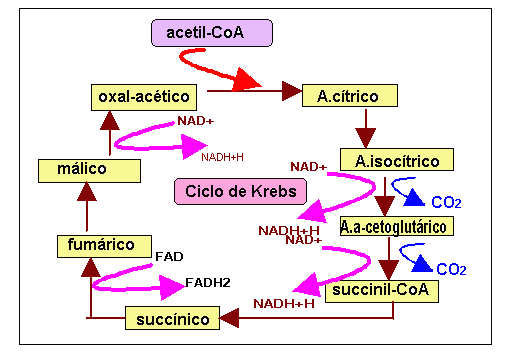
El ciclo de Krebs
Ocurre en las mitocondrias de las células eucariotas y en el citoplasma de las células procariotas.
El catabolismo glucídico y lipídico (a través de la glucolisis y la beta oxidación), produce acetil-CoA, un grupo acetilo enlazado al coenzima A. El acetil-CoA constituye el principal sustrato del ciclo. Su entrada consiste en una condensación con oxalacetato, al generar citrato. Al término del ciclo mismo, los dos átomos de carbono introducidos por el acetil-CoA serán oxidados en dos moléculas de CO2, regenerando de nuevo oxalacetato capaz de condensar con acetil-CoA. La producción relevante desde el punto de vista energético, sin embargo, es a partir de una molécula de GTP (utilizada inmediatamente para regenerar una molécula d e ATP), de tres moléculas de NADH y una de FADH2.
Los cofactores reducidos, NADH y FADH2, se comportan como intermediarios óxido/reductores. Cuando están reducidos, son capaces de transportar electrones a energía relativamente alta (por ejemplo sustraída a los sustratos oxidados en la glucolisis o en el mismo ciclo de Krebs), hasta la cadena respiratoria mitocondrial. Cerca de tal cadena, ese reoxidan a NAD+ y a FAD, y ceden los electrones a la cadena misma, que será así capaz de regenerar moléculas de ADP y ATP.
La reacción neta es la siguiente:
Acetil-CoA + 3 NAD+ + FAD + ADP + Pi => CoA-SH + 3 NADH + H+ + FADH2 + ATP + 2 CO2
La energía que se saca de la ruptura completa de una molécula de glucosa pasa los tres estadios de la respiración celular (glucolisis, ciclo de Krebs y cadena de transporte de electrones), es idealmente de 36 moléculas de ATP. En realidad son 38 las moléculas netas de ATP que se producen, pero dos de ellas se consumen para transportar ( mediante transporte activo), desde el citoplasma a la matriz mitocondrial las dos moléculas de NADH + H+ producidas en la glucolisis.
Etapas del Ciclo de Krebs
Etapas del Ciclo de Krebs
Reacción 1: Citrato sintasa (De oxalacetato a citrato)
El sitio activo de la enzima, activa el acetil-CoA para hacerlo afín a un centro carbonoso del oxalacetato. Como consecuencia de la unión entre las dos moléculas, el grupo tioéster (CoA) se hidroliza, formando así la molécula de citrato.
La reacción es sumamente exoergónica (ΔG'°=-31.4 kJ/mol), motivo por el cual este paso es irreversible. El citrato producido por la enzima, además, es capaz de inhibir competitivamente la actividad de la enzima. Incluso estando la reacción muy favorecida, porque es exoergónica, la citrato sintasa puede ser perfectamente regulada. Este aspecto tiene una notable importancia biológica, puesto que permite una completa regulación del ciclo de Krebs completo, convirtiendo a la enzima en una especie de marcapasos del ciclo.
Reacción 2: Aconitasa (De citrato a isocitrato)
La aconitasa cataliza la isomerización del citrato a isocitrato, por la formación de cis-aconitato. La enzima cataliza también la reacción inversa, pero en el ciclo de Krebs tal reacción es unidireccional a causa de la ley de acción de masa: las concentraciones (en condiciones estándar) de citrato (91%), del intermediario cis-aconitato (3%) y de isocitrato (6%), empujan decididamente la reacción hacia la producción de isocitrato.
En el sitio activo de la enzima está presente un clúster hierro-azufre que, junto a algunos residuos de aminoácidos polares, liga el sustrato. En concreto, la unión al sustrato se asegura por la presencia de un resto de serina, de arginina, de histidina y de aspartato, que permiten sólo la unión estereospecifica del citrato 1R,2S, rechazando la forma opuesta.
Reacción 3: Isocitrato deshidrogenasa (De isocitrato a oxoglutarato)
La isocitrato deshidrogenasa mitocondrial es una enzima dependiente de la presencia de NAD+ y de Mn2+ o Mg2+. Inicialmente, la enzima cataliza la oxidación del isocitrato a oxalsuccinato, lo que genera una molécula de NADH a partir de NAD+. Sucesivamente, la presencia de un ión bivalente, que forma un complejo con los oxígenos del grupo carboxilo en posición alfa, aumenta la electronegatividad de esa región molecular. Esto genera una reorganización de los electrones en la molécula, con la consiguiente rotura de la unión entre el carbono en posición gamma y el grupo carboxilo adyacente. De este modo se tiene una descarboxilación, es decir, la salida de una molécula de CO2, que conduce a la formación de α-cetoglutarato, caracteriz ado por dos carboxilos en las extremidades y una cetona en posición alfa con respecto de uno de los dos grupos carboxilo.
Reacción 4: α-cetoglutarato deshidrogenasa (De oxoglutarato a Succinil-CoA)
Después de la conversión del isocitrato en α-cetoglutarato se produce una segunda reacción de descarboxilación oxidativa, que lleva a la formación de succinil CoA. La descarboxilación oxidativa del α-chetoglutarato es muy parecida a la del piruvato, otro α-cetoácido.
Ambas reacciones incluyen la descarboxilación de un α-cetoácido y la consiguiente producción de una unión tioéster a alta energía con la coenzima A. Los complejos que catalizan tales reacciones son parecidos entre ellos.
La α-cetoglutarato deshidrogenasa (o, más correctamente, oxoglutarato deshidrogenasa), está compuesta de tres enzimas diferentes:
* Subunidad E1: las dos cetoglutarato deshidrogenasas.
* Subunidad E2: la transuccinilasa.
(La subunidad E1 y E2 presentan una gran homología con las de la piruvato deshidrogenasa.)
* Subunidad E3: la dihidrolipoamida deshidrogenasa, que es el mismo polipéptido presente en el otro complejo enzimático.
* Subunidad E1: las dos cetoglutarato deshidrogenasas.
* Subunidad E2: la transuccinilasa.
(La subunidad E1 y E2 presentan una gran homología con las de la piruvato deshidrogenasa.)
* Subunidad E3: la dihidrolipoamida deshidrogenasa, que es el mismo polipéptido presente en el otro complejo enzimático.
Reacción 5: Succinil-CoA sintetasa (De Succinil-CoA a succinato)
El succinil-CoA es un tioéster a alta energía (su ΔG°′ de hidrólisis está en unos -33.5 kJ mol-1 , parecido al del ATP que es de -30.5 kJ mol-1). La citrato sintasa se sirve de un intermediario con tal unión a alta energía para llevar a cabo la fusión entre una molécula con dos átomos de carbono (acetil-CoA) y una con cuatro (oxalacetato). La enzima succinil-CoA sintetasa se sirve de tal energía para fosforilar un nucleósido difosfato purinico como el GDP.
La energía procedente del tioéster viene convertida en energía ligada a una unión fosfato. El primer paso de la reacción genera un nuevo intermediario a alta energía, conocido como succinil fosfato. Sucesivamente, una histidina presente en el sitio catalítico remueve el fosfato de la molécula glucídica, generando el producto succinato y una molécula de fosfohistidina, que dona velozmente el fosfato a un nucleósido difosfato, recargándolo a trifosfato. Se trata del único paso del ciclo de Krebs en el que se prod uce una fosforilación a nivel de sustrato.
El GTP está implicado principalmente en las rutas de transducción de señales, pero su papel en un proceso energético como el ciclo de Krebs es, en cambio, esencialmente trasladar grupos fosfato hacia el ATP, en una reacción catalizada por la enzima nucleósid difosfoquinasa.
Reacción 6: Succinato deshidrogenasa (De succinato a fumarato)
La parte final del ciclo consiste en la reorganización de moléculas a cuatro átomos de carbono hasta la regeneración del oxalacetato. Para que eso sea posible, el grupo metilo presente en el succinato tiene que convertirse en un carbonilo. Como ocurre en otras rutas, por ejem plo en la beta oxidación de los ácidos grasos, tal conversión ocurre mediante tres pasos: una primera oxidación, una hidratación y una segunda oxidación. Estos tres pasos, además de regenerar oxalacetato, permiten la extracción ulterior de energía mediante la formación de FADH2 y NADH.
La primera reacción de oxidación es catalizada por el complejo enzimático de la succinato deshidrogenasa, la única enzima del ciclo que tiene como aceptor de hidrógeno al FAD en vez de al NAD+. El FAD es enlazado de modo covalente a la enzima por un residuo de histidina. La enzima se vale del FAD ya que la energía asociada a la reacción no es suficiente para reducir el NAD+.
El complejo enzimático también es el único del ciclo que pasa dentro de la membrana mitocondrial. Tal posición se debe a la i mplicación de la enzima en la cadena de transporte de los electrones. Los electrones pasados sobre el FAD se introducen directamente en la cadena gracias a la unión estable entre la enzima y el cofactor mismo.
Reacción 7: Fumarasa (De fumarato a L-malato)
La fumarasa cataliza la adición en trans de un protón y un grupo OH- procedentes de una molécula de agua. La hidratación del fumarato produce L-malato.
Reacción 8: Malato deshidrogenasa (De L-malato a oxalacetato)
La última reacc ión del ciclo de Krebs consiste en la oxidación del malato a oxalacetato. La reacción, catalizada por la malato deshidrogenasa, utiliza otra molécula de NAD+ como aceptor de hidrógeno, produciendo NADH.
La energía libre de Gibbs asociada con esta última reacción es decididamente positiva, a diferencia de las otras del ciclo. La actividad de la enzima es remolcada por el consumo de oxalacetato por parte de la citrato sintasa, y de NADH por parte de la cadena de transporte de electrones.
LA CADENA DE TRANSPORTE DE ELECTRONES:

FOSFORILACIÓN OXIDATIVA:
Es cuando los protones que fluyen por un canal interior de las ATP-sintetasas provocan cambios en tres lugares y producen la unión de un ADP con un P generando ATP.

BIBLIOGRAFIA:
{kind=link}
ai weee no ma t kedo muy xido spro i sakes l 10 jajajajajaja
ResponderEliminar